Семейство ильмовых объединяет две довольно обособленные группы древесных растений, различающиеся по строению цветков, пыльцевых зерен, плодов, зародышей, анатомии листьев, основному хромосомному числу, составу химических веществ и пр. Этим двум группам обычно придают ранг подсемейств или реже самостоятельных семейств. В данном издании мы принимаем их как подсемейства ильмовые (Ulmoideae) и каркасовые (Celtidoideae).
В семействе ильмовых только древесные растения с простыми очередными листьями и быстро опадающими прилистниками. Их пазушные соцветия объединяют мелкие ветроопыляемые цветки с простым чашечковидным околоцветником, расчлененным на 4 — 5 (9) лопастей; им противостоит примерно такое же число тычинок. Завязь верхняя, одногнездная, развивающаяся в односемянный невскрывающийся плод.
Подсемейство ильмовых — небольшая однородная группа, наиболее обособленная и наиболее примитивная в порядке крапивных. Оно объединяет 6 родов, в которые входит около 50 видов древесных растений. Центральное место среди ильмовых принадлежит роду ильм (Ulmus), включающему более 75% видов подсемейства, распространенных от умеренной до тропической зоны северного полушария.
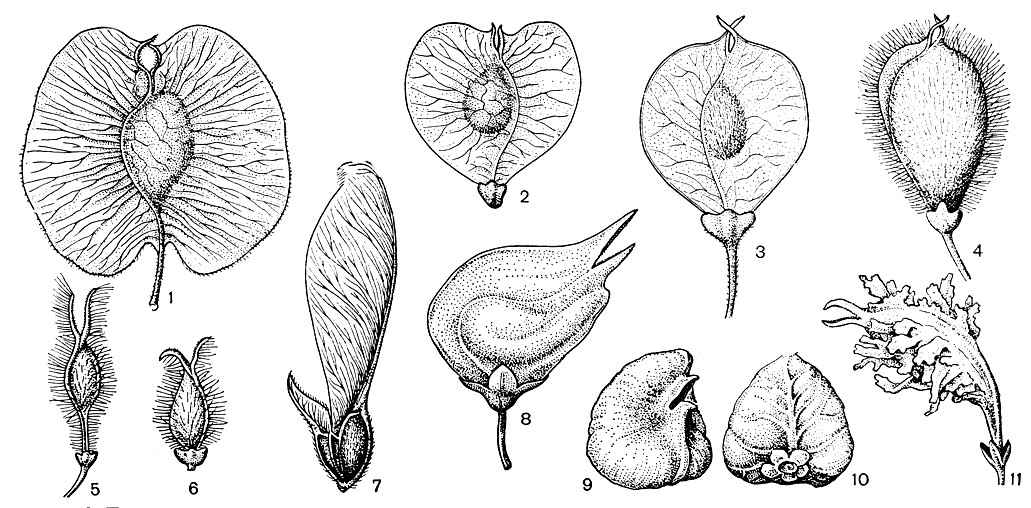 Рис. 1. Плоды ильмовых: 1 — голоптелея цельнолистная (Holoptelea integrifolia); 2 — ильм мелколистный (Ulmus pumila); 3 — ильм Валлиха (U. wallichiana); 4 — ильм Томаса (U. thomasii); 5 — ильм мексиканский (U. mexicana); 6 — ильм мохнатый (U. villosa); 7 — филлостилон бразильский (Phyllostylon brasiliense); 8 — гемиптелея Давида (Hemiptelea davidii); 9 и 10 — дзельква граболистная (Zelkova carpinifolia); 11 — планера водная (Planera aquatica)
Рис. 1. Плоды ильмовых: 1 — голоптелея цельнолистная (Holoptelea integrifolia); 2 — ильм мелколистный (Ulmus pumila); 3 — ильм Валлиха (U. wallichiana); 4 — ильм Томаса (U. thomasii); 5 — ильм мексиканский (U. mexicana); 6 — ильм мохнатый (U. villosa); 7 — филлостилон бразильский (Phyllostylon brasiliense); 8 — гемиптелея Давида (Hemiptelea davidii); 9 и 10 — дзельква граболистная (Zelkova carpinifolia); 11 — планера водная (Planera aquatica)У всех родов подсемейства ильмовых, даже настоящих тропических, молодые формирующиеся побеги прикрыты почечными чешуями. Цветки обоеполые или обоеполые и мужские, нити тычинок в почках прямые. В порядке крапивных только у ильмовых четко заметно, что их гинецей образован двумя сросшимися плодолистиками: в одном из них развивается семязачаток, другой редуцирован и остается стерильным. Верхние части плодолистиков не срастаются и несут на внутренней стороне рыльцевые поверхности. Столбика нет. Завязь одногнездная, сплюснутая.
Простота строения цветка ильмовых, как и всего порядка крапивных, является вторичной. Его упрощение шло за счет редукции, слияния и утраты отдельных органов (части тычинок, венчика, долей чашечки и плодолистиков). Об этом можно судить по сохранности у примитивных групп видов большого числа частей цветка, а также по остаткам сосудов проводящей системы уже несуществующих органов. Редукционные процессы в цветках начались, вероятно, очень давно. Во всяком случае, судя по ископаемым отпечаткам, цветки ильмовых, возраст которых определяют в 20 — 30 млн. лет, уже имели строение, аналогичное современному.
У всех родов ильмовых завязи сходны по строению, однако превращение их в плоды проходит крайне специфично у каждого рода, в результате чего и сами плоды резко различаются по форме и строению (рис. 1). Плоды наших ильмов и голоптелей (Holoptelea) — крылатые семянки, но у некоторых ильмов развиваются бескрылые плоды. Остальным родам ильмовых свойственны орехи: у филло-стилона бразильского (Phyllostylon brasiliense) они оканчиваются двумя узкими неодинаковыми крыльями с ребрами по внешнему краю и с рыльцами по внутреннему; у гемиптелеи Давида (Hemiptelea davidii) орехи косые, однокрылые, вздутые, горбатые; орехи у видов дзелъквы (Zelkova) не имеют крыльев, а у планеры водной (Planera aquatica) вместо крыльев развиты полумясистые гребневидные выросты. Плоды хорошо приспособлены к переносу ветром. Этому способствуют их крылья, крупные воздухоносные полости в межклетниках околоплодника, небольшая масса и уплощенная форма плода, нередко окаймленного ресничками, или увеличенная полость гнезда семени. Зоохория не имеет здесь большого значения, хотя животные охотно поедают плоды ильмовых.
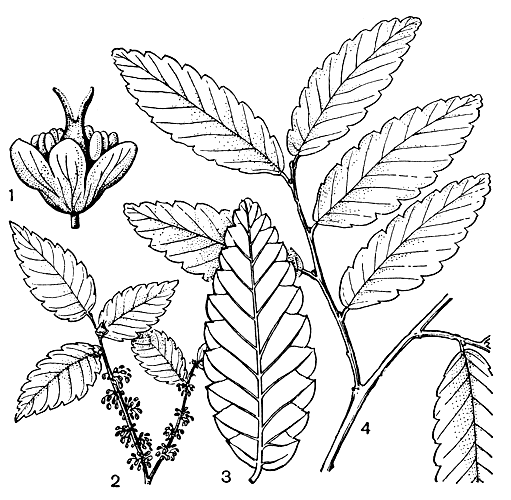 Рис. 2. Дзельква граболистная (Zelkova carpinifolia): 1 — цветок; 2 — цветоносные побеги; 3 — лист (нижняя сторона); 4 — вегетативный побег
Рис. 2. Дзельква граболистная (Zelkova carpinifolia): 1 — цветок; 2 — цветоносные побеги; 3 — лист (нижняя сторона); 4 — вегетативный побегВ умеренном климате, с резкой сменой времен года, ильмовые — обычно листопадные летнезеленые растения, в субтропической и особенно тропической зоне среди них появляются полулистопадные, реже вечнозеленые формы. Тропические полулистопадные голоптелея и филлостилон обычно сбрасывают листья перед цветением, но продолжительность пребывания в безлистном состоянии у них резко колеблется по годам и бывает связана с условиями жизни и с возрастом дерева. На Кубе в отдельные годы молодые растения филлостилона сохраняют большую часть листвы круглый год, а в засушливые годы взрослые деревья стоят без листьев около 3 месяцев. Дзельква и гемиптелея — листопадные растения, а род ильм представлен всем разнообразием форм, и с севера на юг к его листопадным видам прибавляются полулистопадные, а в тропиках также вечнозеленые.
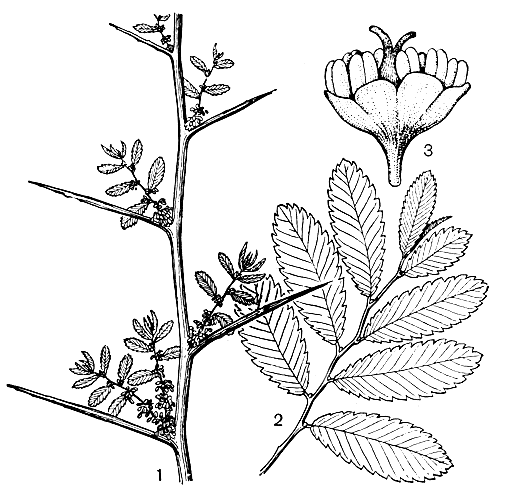 Рис. 3. Гемиптелея Давида (Hemiptelea davidii): 1 — ветвь с колючками и цветоносными побегами; 2 — вегетативный побег; 3 — цветок
Рис. 3. Гемиптелея Давида (Hemiptelea davidii): 1 — ветвь с колючками и цветоносными побегами; 2 — вегетативный побег; 3 — цветокЕсли проследить сезонное развитие видов ильма от умеренной до тропической зоны, можно подметить очень любопытные закономерности не только в характере листопадности, но и в ритмике цветения. У ильмов умеренной зоны цветки сформированы в почках уже в начале лета, однако зацветают они лишь на следующий год и, таким образом, находятся в почках примерно 10 месяцев. Цветут ильмы ранней весной еще в безлистном состоянии. Южнее, в Средней Азии и Средиземноморье, ильмы зацветают в феврале и даже в январе, и период нахождения цветков в почках сокращается примерно до 7 месяцев. В субтропических районах Америки, Японии и Китая есть полулистопадные ильмы, цветущие осенью того же года. Их цветки находятся в почках лишь 3 — 4 месяца, соцветия появляются осенью, когда многие листья еще не опали. Часть соцветий появляется в пазухах этих листьев, что создает впечатление олиственности цветоносных побегов. Наконец еще южнее, в тропиках Юго-Восточной Азии вечнозеленый ильм ланцетолистный (Ulmus lanceifolia) цветет в начале лета, по-видимому, сразу же после окончания формирования цветков; время нахождения в почках его цветков сведено до минимума.
Соцветия ильмов, голоптелеи, филлостилона и планеры безлистны и формируются в специализированных почках, как правило, не несущих зачатков настоящих листьев. Напротив, цветоносные побеги видов дзельквы (рис. 2) и гемиптелеи (рис. 3) не имеют строгой специализации. Их цветки формируются вместе с листьями в одних и тех же почках, зацветают весной — в начале лета, сразу же после развертывания листьев; плоды созревают лишь в конце лета или осенью. У ильмов же, как и у планеры, формирование плодов длится около месяца и уже в конце весны — начале лета плоды созревают и опадают.
Семена ильмов имеют плоский, прямой зародыш, защищенный трехслойной семенной кожурой (четвертый внутренний однорядный слой образован клетками эндосперма) и четы-рехслойным пленчатым околоплодником. На влажном субстрате семена прорастают через несколько суток без периода покоя. Развившиеся всходы резко отличаются от побегов взрослых растений. Это явление, известное у многих растений, получило название гетеро-бластного развития. У ильмов оно заключается в том, что обычные побеги их имеют билатерально-симметричное строение: пластинки листьев асимметричны, прилистники неодинаковы по форме и размеру, листорасположение двурядно-очередное. На побегах никогда не формируется верхушечная почка, и после остановки роста побега его верхняя часть отмирает. В отличие от этого у всходов ильмов главный побег радиально-симметричный: пластинки его листьев более или менее симметричны, прилистники одинаковы, листья расположены на побеге накрест супротивно. На вершине такого побега формируется терминальная почка. Правда, специальных почечных чешуи при этом не возникает, и защиту верхушечного конуса нарастания принимают на себя прилистники верхних листьев, как это свойственно многим тропическим растениям, не образующим почечных чешуи. Эти прилистники-чешуи сохраняются на побеге до следующей весны, т. е. живут гораздо дольше, чем листья, в то время как обычные прилистники у ильмов опадают значительно раньше листьев — в начале лета.
Своеобразие главного побега всхода еще и в том, что он развивается по моноподиальному типу и формируется верхушечной (по положению) меристемой. Все последующие побеги, в том числе и продолжающие главную ось растения (ствол), возникают за счет деятельности пазушной (боковой) меристемы; верхушечная меристема у ильмов обычно вскоре после образования терминальной почки всхода отмирает. Побег, образовавшийся из верхней боковой почки, растет наиболее быстро и, перевершинивая материнский побег, становится осевым. Это из года в год повторяющееся перевершинивание — характерная особенность роста ствола и ветвей ильмовых, позволяющая относить их к типичным симподиальным растениям.
Побеги некоторых ильмов привлекают внимание своеобразными пробковыми наростами, особенно характерными у молодых растений, растущих на сухих, хорошо освещенных местах. Выросты совсем другого типа возникают иногда на стволах старых деревьев вяза мелколистного в ксерофитных формациях Восточного Казахстана. Это капы — огромные наплывы уплотненной, необычайно крепкой древесины. Интересное видоизменение укороченных побегов, превращенных в настоящие колючки, характерно для гемиптелеи (рис. 3) — единственного колючего деревца среди ильмовых.
Листья ильмов даже на одном побеге могут резко различаться размером и очертаниями. Это позволяет им наиболее выгодно располагаться по отношению к свету — в одной плоскости — в виде сплошного мозаичного покрова (листовая мозаика). Жилкование листьев ильмовых типичное перистое, краебежное, с мощной средней жилкой и короткими боковыми, обычно оканчивающимися в зубцах листа. Поверхность листьев нередко опушена мягкими или грубыми волосками, а у некоторых видов ильмов листья снизу покрыты мельчайшими железистыми волосками: точечными (у наших берестов) или палочковидными (у гималайского илъма мохнатого — U. villosa). Цвет этих волосков-железок меняется по мере старения листа: от бесцветных у молодых развивающихся листьев, оранжевых в начале лета, красных летом до почти черных осенью.
Корневая система у ильмовых мощная, с отдельными вглубь идущими корнями и массой поверхностных. У крупных деревьев иногда возникают досковидные корни, выполняющие опорную функцию и столь характерные для деревьев тропических дождевых лесов. Эти корни достигают в высоту 1,5 м в месте отхождения от стволов у тропических голоптелей. У ильмов умеренной зоны (вяза гладкого — U. laevis, илъма долинного — U. japonica) их высота обычно 30 — 50 см, но тип их строения такой же, как у тропических деревьев. Впрочем, по данным И. В. Грушвицкого (1955), отдельные деревья ильма долинного на юге Приморского края имеют полутора- и даже двухметровые досковидные корни.
Представители всех исследованных родов ильмовых — микоризообразующие растения, особенно обильная микориза бывает на корнях береста, которые нередко покрываются своеобразными микоризными чехлами.
Корневая система имеет большое значение в вегетативном размножении ильмовых благодаря образованию корневых отпрысков. У гексаплоидной гемиптелеи Давида, образующей много бессемянных плодов, корнеотпрысковый тип возобновления нередко преобладает над семенным. Часто размножается корнеотпрысками и берест.
Сведения о продолжительности жизни ильмовых противоречивы, но достоверно известны ильмы и дзельквы, дожившие до 500 лет (возраст отдельных деревьев дзельквы граболистной — Zelkova carpinifolia — в Талыше определяют в 800 — 850 лет). Такие долгожители нередко достигают и максимальных для этих видов размеров: в высоту до 35 — 40 м и в диаметре 3 — 4 м. Ильмы-великаны сохранились также в приречных лесах Дальнего Востока (ильм долинный), в тропических лесах Мексики (ильм мексиканский — Ulmus mexicana) и Юго-Восточной Азии (ильм ланцетолистыый); больших размеров достигает голоптелея цельнолистная (Holoptelea integrifolia) в тропических лесах Индии. Представители других родов ильмовых — небольшие деревья, высотой 4 — 18 м.
Современный ареал подсемейства ильмовых охватывает огромную территорию, в пределах которой большинство родов имеет разорванное (дизъюнктивное) распространение, и появление этих дизъюнкций обычно связывают с третичным или верхнемеловым временем.
Палеоботанические данные указывают, что в миоцене в умеренных и теплоумеренных флорах третичной Бореальной области, соответствующей территориям современных Евразии и Северной Америки, ильмовые были широко распространены и представлены большим разнообразием форм (роды ильм, дзельква, планера). Разрывы современных ареалов тропических родов голоптелеи (Западная Индия — экваториальная Африка) и филлостилона (Бразилия — Карибская флористическая область) подтверждают древность и этих родов и позволяют предполагать более широкое распространение их в прошлом. Даже монотипный род гемиптелея, современный ареал которого ограничен небольшим районом в Восточной Азии, в миоцене — плиоцене был распространен в Средней и Южной Европе.
Планера водная — единственный современный представитель рода планера — реликтовое растение, изредка встречающееся во Флориде и прилегающих районах юго-востока Северной Америки. Она произрастает в древних лесах с господством болотного кипариса (Taxodium distichum), почва в которых большую часть года покрыта водой.
Явно реликтовым является современный ареал дзельквы (Япония — районы Южного и Центрального Китая — Закавказье — районы Передней Азии — остров Крит).
Среди ильмовых только род ильм имеет неразорванный (сплошной) обширный ареал, но в пределах его есть любопытные дизъюнкции в распространении близких видов одной секции. Так, наш европейский вяз гладкий (Ulmus laevis) трансатлантической дизъюнкцией отделен от близкородственного вяза американского (U. americana). Морфологически эти виды почти неотличимы, но имеют разную степень плоидности: европейский — диплоид (2п = 28), американский — тетраплоид (2п = 56). Аналогична дизъюнкция и между диплоидными видами: европейским ильмом горным (U. glabra) и американским ильмом красным (U. rubra).
В нашей стране представители рода ильм известны под названиями ильмы, вязы, бересты, карагачи. Их узнают обычно по двоякозубчатым неравнобоким в основании листьям и плодам-крылаткам, появляющимся в начале лета.
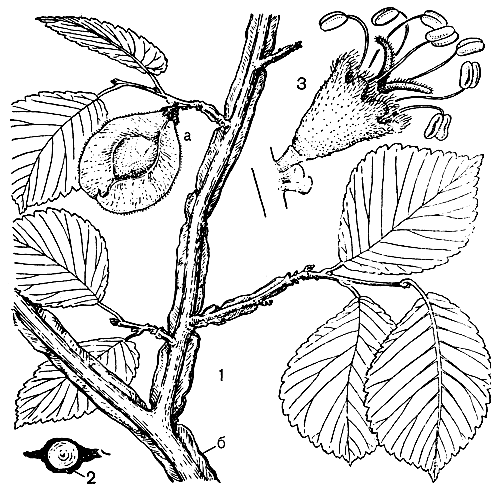 Рис. 4. Ильм крупноплодный (Ulmus macrocarpa): 1 — ветвь с плодом (а) и пробковым наростом (б); 2 — поперечный срез ветви (пробковый нарост затушеван); 3 — цветок
Рис. 4. Ильм крупноплодный (Ulmus macrocarpa): 1 — ветвь с плодом (а) и пробковым наростом (б); 2 — поперечный срез ветви (пробковый нарост затушеван); 3 — цветокВ широколиственных лесах Европейской части СССР наиболее обычны вяз гладкий и ильм горный — крупные деревья, достигающие в высоту 25 — 27 м. Оба вида имеют большой широтный диапазон распространения, особенно велик он у вяза гладкого. Простираясь от берегов Онежского озера до пустынь Прикаспия, его ареал охватывает зону полупустынь, степей, лесостепей, широколиственных лесов и темно-хвойной тайги.
В лесостепной зоне более обычен берест (U. campestris) — сравнительно небольшое опушечное дерево, нередко привлекающее внимание пробковыми наростами на ветвях.
По долинам рек Дальнего Востока в широколиственных и кедрово-широколиственных лесах нередки огромные белокорые ильмы долинные, здесь же растут небольшие деревья ильма лопастного (U. laciniata), более обычного в горных лесах Приморья. На Дальнем Востоке и в Забайкалье распространены также ильм крупноплодный (U. macrocarpa, рис. 4) и вяз мелколистный — пионерные породы открытых местообитаний, иногда образующие ксерофитные редколесья.
Ильм крупноплодный — небольшое деревце, на скалах и осыпях нередко развивающийся как кустарниковидное растение, обильно плодоносящее даже при высоте 50-70 см. Его крылатки наиболее крупные (диаметром до 3 — 4 см), пробковые наросты на молодых побегах растут в одной плоскости, отчего побеги также выглядят крылатыми.
Огромное значение в озеленении и защитных насаждениях в аридных странах почти всех континентов имеет вяз мелколистный (U. pumilа). Естественный ареал его простирается от гор Западного Тянь-Шаня через пустыни Монголии и Китая до Забайкалья и Дальнего Востока. В пустынях Гоби он нередко оказывается единственной древесной породой. Здесь это низкорослое дерево (высотой 2 — 6 м) с небольшой кроной и мощным стволом, диаметром до 1 — 1,5 м. В Восточном Казахстане, в бассейне реки Или, столетние вязы имеют высоту 8 — 12 м, такой же высоты они достигают в поймах рек Дальнего Востока и Забайкалья, но в культуре, особенно вдоль арыков в Средней Азии, они могут превышать в высоту 25 м и развивать мощную раскидистую крону.
С начала разведения этого вида за пределами естественного ареала не прошло еще и ста лет, но сейчас уже его «культурный» ареал опоясывает все северное полушарие, захватывая и отдельные участки южного (районы Австралии и Аргентины). В районе Великих равнин (Северная Америка) вяз мелколистный ведет себя как абориген и включен в местные флоры. В нашей стране он — излюбленная порода в озеленении южных городов и поселков на всем протяжении от восточных до западных границ.
Представители рода ильм существуют на Земле десятки миллионов лет и, судя по ископаемым находкам, не изменились сколько-нибудь существенно за этот период времени, несмотря на неоднократную и подчас резкую смену условий жизни. Это говорит об огромном приспособительном (адаптивном) потенциале, который виден и в настоящее время по широте экологии и современному распространению рода. Являясь характерными компонентами широколиственных лесов, ильмы растут также в пустынях и за полярным кругом, вдоль пересыхающих рек (уэдов) в Северной Африке и близ экватора, на Суматре и Сулавеси, в тропических лесах Юньнани, в горах Мексики и в Гималаях.
Ильмы — неприхотливые растения, переносящие недостаток влаги и избыточное проточное увлажнение, они способны расти на засоленных почвах, каменистых россыпях и скалах, на приречных песках и галечниках, мириться с недостатком тепла на севере и избытком его в жарких пустынях, с колебанием уровня воды рек и озер, по берегам которых эти деревья встречаются наиболее часто. И именно участки с крайне изменчивыми факторами среды, в той или иной степени неблагоприятные для развития леса и роста деревьев вообще, наиболее обычны для ценозов с преобладанием ильмов.
В равнинных широколиственных лесах, оптимальных для развития большинства наших ильмов, они встречаются лишь в небольшой примеси к основным породам, прочно занимая место пород-ассектаторов (дополнительных). Даже в поймах крупных рек, где ильмы нередко формируют участки чистых насаждений, их произрастание обычно связано с узкой полосой, отличающейся наиболее изменчивым водным режимом, на стыке пойменных дубрав и зарослей ив или ольхи. В сухие годы эта полоса неблагоприятна для развития ив, во влажные — для дуба.
Ильмовые издавна используют для различных целей. Слизистые выделения луба ильмовых обладают бактерицидными свойствами, их, как и семена, применяют в народной медицине. Из семян ильмов получают также ценное техническое масло. Недозрелые плоды вяза мелколистного в Китае употребляют в пищу как салат.
В ряде горных районов Азии и Закавказья ветви ильмов и дзельквы заготовляют на корм скоту. В ландшафтах этих горных стран и особенно в Гималаях не редкость изуродованные деревья ильмов, ветви которых обрублены почти до вершины ствола.
Большую хозяйственную ценность имеет древесина ильмовых. Уже в ископаемых поселениях человека в Европе были найдены дома, построенные из ильмов. В прошлом веке древесину ильмов и дзельквы широко использовали как строительный материал, особенно для построек в воде: на сваи, в кораблестроении и пр. Идет она также на производство мебели и фанеры.
В настоящее время, когда запасы древесины ильмовых в естественных насаждениях значительно убавились, наибольшую пользу ильмы приносят как породы-озеленители и неизменные компоненты защитных насаждений. Быстрота роста ильмов, их декоративность, нетребовательность к почвенному питанию, способность выдерживать недостаток влаги и сильные ветры, значительные колебания температуры и задымленность воздуха с давних пор сделали их излюбленными деревьями в озеленении городов в странах северного полушария.
Ильмы на улицах, в садах и парках культивируются по всей Евразии (вяз мелколистный, вяз гладкий, ильм горный, ильм долинный, берест), в Африке (вяз мелколистный, ильм седоватый), в Северной Америке (вяз американский, вяз мелколистный, ильм Томаса, ильм красный). Помимо обычных видов, в озеленении используют также ряд своеобразных декоративных форм, закрепленных в культуре. Это плакучие и пирамидальные ильмы, а также знаменитые густокронные карагачи — вяз густой (U. densa) и вяз Андросова (U. androssowii), украшающие улицы, сады и парки республик Средней Азии, отдельных районов Закавказья и Передней Азии. Их необычайно густая шаровидная или продолговатая крона почти не пропускает солнечных лучей и укрывает от солнца в любое время дня, что придает им исключительную ценность в районах жарких пустынь. Густокронные виды отличаются очень медленным ростом, и поэтому их, как правило, прививают на обычные деревца береста или вяза мелколистного.
За последние 60 лет среди ильмовых распространилось заболевание, называемое по месту его открытия голландской болезнью. Ей подвержены все виды ильмов (устойчив лишь вяз мелколистный). Наиболее действенной мерой, препятствующей развитию болезни, является инъекция антибиотиков в ствол растения.
Подсемейство каркасовых — вечнозеленые, полулистопадные или листопадные деревья, реже вечнозеленые лазящие лианы, распространенные в тропиках и субтропиках всех частей света. Из 9 составляющих подсемейство родов (около 80 видов) только один монотипный род птероцелътис (Pteroceltis) не выходит за пределы теплоумеренной зоны, остальные 8 родов включают преимущественно тропические растения, и лишь отдельные их виды произрастают в районах умеренно теплого климата.
По большинству морфологических признаков и общему уровню развития каркасовые являются значительно более специализированной группой, чем ильмовые.
В подсемействе преобладают виды с однополыми цветками, хотя у некоторых обычен небольшой процент обоеполых цветков. В тепло-умеренной зоне каркасовые представлены только однодомными растениями, в тропиках — и однодомными и двудомными, причем даже в пределах одного вида можно видеть разную степень дифференцировки деревьев по преобладанию женских или мужских цветков. Хетакма остистая (Ghaetacme aristata), например, в тропиках Восточной Африки почти всегда имеет двудомные цветки, а в Южной Африке чаще развивается как однодомное растение. Все переходы от однодомности к двудомности отмечены у тремы восточной (Trema orientalis), тремы Ламарка (Т. lamarckiana), каркаса Дюрана (Celtis durandii) и др.
Мужские цветки каркасовых собраны в многоцветковые соцветия в пазухах чешуевидных листьев, женские расположены выше по побегу в пазухах зеленых листьев, по 1 — 3, или в сложных многоцветковых соцветиях.
В отличие от подсемейства ильмовых, характеризующихся сухими плодами, все каркасовые имеют один тип плода — костянку, но строение ее, размеры и форма разнообразны (рис. 5).
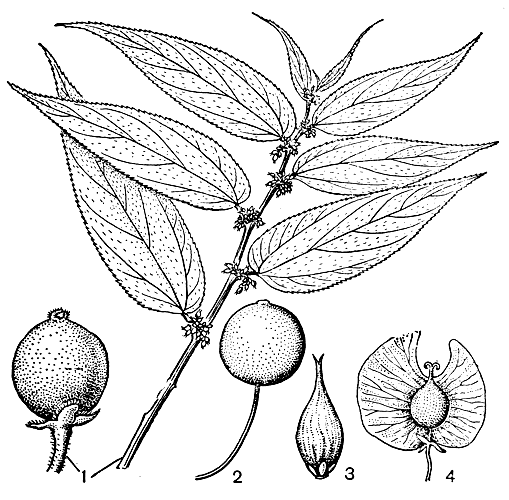 Рис. 5. Плоды каркасовых: 1 — трема мелкоцветковая (Trema micrantha), плод и ветвь с плодами; 2 — каркас кавказский (Geltis caucasica); 3 — афананта длинноостроконечная (Aphananthe cuspidata); 4 — птероцельтис Татаринова (Pteroceltis tatarinovii)
Рис. 5. Плоды каркасовых: 1 — трема мелкоцветковая (Trema micrantha), плод и ветвь с плодами; 2 — каркас кавказский (Geltis caucasica); 3 — афананта длинноостроконечная (Aphananthe cuspidata); 4 — птероцельтис Татаринова (Pteroceltis tatarinovii)Семена каркасовых обычно округлые, зародыш поперек согнут, поперек сложен или свернут в спираль; в зрелых семенах эндосперм сохраняется, он окружает зародыш и заполняет углубления его складок; семенная кожура всегда однослойная, околоплодник 3 — 4-слойный.
Основное хромосомное число у каркасовых такое же, как у коноплевых (х=10). Характерна полиплоидия, причем отдельные виды достигают очень высокой степени плоидности (трема амбоинская — Trema amboinensis — 16-плоид!).
Листья у большинства каркасовых имеют 3 четко выраженные базальные жилки, что приближает их жилкование к пальчатому типу. У большинства каркасов Южной Америки на нижней стороне листовой пластинки в результате разрастания оснований этих жилок (вероятно, под воздействием поселяющихся в них клещиков) развиваются специфические структуры, похожие на вздутые кармашки. Наконец, листьям каркасовых свойственны округлые цистолиты и многообразный характер опушения (у вестиндской тремы Ламарка на одном листе развивается до 4 типов волосков, включая и железистые).
Основными родами подсемейства каркасовых являются 2 пантропических рода: каркас и трема (Trema), включающие более 85% всех видов. Каркас — не только самый крупный (более 50 видов), наиболее полиморфный, но и наиболее широко распространенный род. Его ареал опоясывает земной шар огромной полосой, северная граница которой колеблется около 40° северной широты, проходя по Японии, континентальной Азии, Кавказу, Южной Европе и Северной Америке; южная идет примерно по 35° южной широты через Новую Каледонию, Восточную Австралию, Капскую область Африки и Южную Аргентину. Несмотря на большую полиморфность, каркасы сохраняют в пределах всего ареала единый тип строения цветков, плодов и листьев. Виды каркаса или диплоидные (число хромосом в соматических клетках 2n = 20), или тетраплоидные (2n = 40) растения. К последним относятся все виды СССР, распространенные также в Средиземноморье: каркас кавказский (С. caucasica), каркас голый (С. glabrata), каркас южный (С. australis), каркас Турнефора (С. tournefortii). Это листопадные, летнезеленые деревья с гладкой светло-серой корой и раскидистой кроной, достигающие в высоту иногда 30 м, со стволом диаметром до 3 м, или растущие небольшими деревцами и приобретающие кустарниковидную форму в неблагоприятных условиях среды (каркас голый и каркас Турнефора).
Виды каркаса занимают различные экологические ниши и растут в разнообразных сообществах. Каркас кавказский и каркас Турнефора чаще распространены в сухих лесах и аридных редколесьях, на низменностях и в горах, нередко поднимаясь до высоты 2500 — 2800 м над уровнем моря, обычно в местах, удаленных от моря. Напротив, каркас южный и каркас голый растут преимущественно в приморских районах. Все эти виды обитают на открытых каменистых склонах и в ущельях, среди скал, на осыпях, по каменистым берегам небольших рек или по склонам к морю.
Богато представлены и разнообразны каркасы тропических стран, где наряду с листопадными и полулистопадными обычны и вечнозеленые растения, причем у многих из них почечные чешуи не развиваются и зачатки формирующегося побега прикрыты лишь прилистниками кроющего листа.
Очень своеобразны каркасы Австралии и особенно Новой Каледонии, отличающиеся цельно-крайними толстыми суккулентными листьями. У каркаса скученного (С. conferta), произрастающего в прибрежной зоне и нередкого среди мангровых зарослей, листья собраны на верхушке побега и иногда супротивны. В тропиках Азии, Африки и Южной Америки каркасы входят в сообщества вечнозеленых дождевых лесов низменностей (каркас Уайта — С. wightii, каркас Милъдбреда — С. mildbraedii), а также горных лесов (каркас Дюрана и др.). Растут тропические каркасы и в сухих вечнозеленых лесах, нередко образуя листопадные редколесья (каркас африканский — С. africana, каркас целънолистный — С. integrifolia).
В нарушенных рубкой лесах тропиков Нового Света интенсивно развиваются вечнозеленые лазящие лианы: каркас игуановый (С. iguanaea), каркас колючий (С. spinosa), каркас боливийский (С. boliviensis). В первые годы жизни эти каркасы растут как прямостоячие деревца или кустарники (высотой 1,5 — 5 м), позднее их верхние ветви интенсивно удлиняются и, цепляясь загнутыми колючками за близ стоящие деревья и кустарники, используют их как опору. Растение таким образом превращается в лазящую лиану и сохраняет этот облик до конца жизни.
В теплоумеренном климате каркасы цветут весной, почти одновременно с распусканием листьев. Их мужские цветки раскрываются на несколько дней раньше обоеполых и женских. Цветки опыляются ветром, и, хотя их посещают насекомые, энтомофилия не имеет большого значения, так как пыльца лишена характерных признаков зерен энтомофильных растений и в момент выпрямления тычиночной нити мгновенно высыпается в окружающее пространство.
Плоды созревают осенью, к этому времени внутренняя часть околоплодника (косточка) становится очень твердой, а мучнистый наружный слой приобретает окраску от ярко-желтой (каркас голый) до почти черной (каркас южный). Костянки каркасов охотно поедаются птицами и ими же распространяются.
Семена каркасов прорастают обычно весной следующего года, всходы, как и у ильмовых, развиваются по гетеробластному типу и в первый же год переходят к симподиальному ветвлению.
Растут каркасы сравнительно медленно и долго живут (до 200, а по отдельным сведениям — до 600 лет). Однако в настоящее время старых крупных деревьев каркаса немного. Высокая ценность его древесины и произрастание главным образом в малолесных районах влекут за собой регулярные рубки молодых деревьев. После рубок деревья сравнительно быстро восстанавливаются благодаря интенсивному порослевому (от пня) возобновлению.
Каркас нередко называют также каменным деревом за его твердую, крепкую, тяжелую (плотность 0,78) древесину. Несмотря на ряд ценных свойств, она все же не имеет большого промышленного значения и идет в настоящее время в основном на мелкие поделки и декоративные изделия. В аридных странах каркасы издавна разводят, любят эти деревья и у нас в Средней Азии, на Кавказе и в Крыму, где часто используют в озеленении поселков и городов, а также в защитных насаждениях.
Наиболее близок к каркасам род птероцельтис, распространенный в Центральном Китае. Единственный вид его был описан русским ботаником К. И. Максимовичем и назван в честь коллектора этого растения — птероцелыпис Татаринова (Pteroceltis tatarinovii). Очень своеобразные крылатые плоды птероцельтиса нередко рассматривают как переходное звено между бескрылыми плодами каркасовых и крылатками ильмовых, что дает основание сближать и сами таксоны, однако сходство плодов только внешнее — это пример конвергентного развития. Плод птероцельтиса — настоящая шаровидная костянка с очень толстым, крепким эндокарпием, выросты которого образуют деревянистые крылья, утончающиеся к краю. Вверху крылья широко расставлены, между ними и изолированно от них, в отличие от ильмовых, расположен столбик с двумя рыльцами (рис. 5). Птероцельтис и по другим признакам типичный представитель подсемейства каркасовых: у него однополые цветки, костянки с 4-слойным околоплодником, однослойная семенная кожура, складчато-сложенный зародыш, 3 жилки в основании листа, основное хромосомное число х равно 20. Растет птероцельтис обычно вдоль рек и по скалистым местам на сравнительно небольших высотах (до 1200 м над уровнем моря). Чаще всего это деревья высотой 12 — 17 м, с раскидистой кроной и короткими толстыми стволами диаметром до 1,5 м.
Очень интересный и, по-видимому, монотипный род хетакма (Ghaetacme) распространен на Мадагаскаре и в экваториальной и Южной Африке. Хетакма остистая развивается как небольшое деревце или кустарник высотой 3 — 7 м, она имеет блестящие кожистые листья, иногда оканчивающиеся тонким заострением — колючкой, однополые цветки и мелкие плоды с твердой косточкой. Вид этот очень полиморфен и включает растения с однодомными и двудомными цветками, представлен сильно опушенными и голыми формами, колючими и без колючек, листья у него зубчатые, колючезубчатые или цельнокрайние. Хетакма остистая произрастает в зоне, переходной от леса к саванне, встречается в листопадных, полулистопадных лесных и кустарниковых формациях, в склерофильных галерейных лесах, где иногда вместе с другими растениями образует непроходимые колючие заросли, столь характерные для этих стран.
В тропических странах широко известен род трема, представленный преимущественно вечнозелеными деревцами, иногда кустарниковидными растениями, высотой 2 — 16 м, распространенными на всех континентах и многих островах от низменностей до 2500 м над уровнем моря. У них негустая распластанная крона, разветвленные пазушные соцветия, несущие многочисленные мелкие, обычно однополые цветки. Плоды — небольшие мясистые костянки, ярко-желто-оранжевые у тремы Ламарка и темные у тремы восточной и тремы мелкоцветковой.
Виды рода трема трудно разграничиваются, и среди систематиков до сих пор имеются разногласия относительно их объема и числа. По-видимому, в роде не более 20 видов, все они близки и образуют полиплоидный ряд с числом хромосом в соматических клетках от 20 до 160.
Тремы — быстро растущие неприхотливые растения, обитающие на опушках вечнозеленых и полулистопадных лесов равнин и горных склонов, обычны вдоль дорог и на вырубках. Виды тремы — обычные компоненты вторичных растительных формаций тропиков, в частности, характерные представители своеобразных вторичных формаций с древовидными папоротниками, развивающихся после пожаров и рубок на месте дождевых горных тропических лесов. На Кубе это сообщество тремы мелкоцветковой (Trema micrantha) и папоротника циатеи древовидной (Cyathea arborea), со сплошным покровом из папоротника орляка хвостатого (Pteridium caudatum). Аналогичные растительные формации характерны и для тропиков Старого Света. На Яве, например, имеются сообщества тремы восточной и циатеи грязноватой (Cyathea contaminans), в подлеске — посконник инулолистный (Eupatorium inulifolium).
Трема пока единственный род в порядке крапивных, у которого обнаружен симбиоз с азот-фиксирующими бактериями. На корнях тремы восточной недавно были найдены клубеньковые бактерии из группы Rhizobium, что дает основание относить ее к почвоулучшаюшим растениям. Возможно, отчасти благодаря этому свойству трему охотно используют на плантациях кофейного дерева и какао для создания негустого полога, под которым и высаживают эти культуры.
Очень близок к роду трема и трудно отличим от него небольшой род параспония (Parasponia), распространенный на островах Океании. Представители обоих родов — типичные пионерныо растения, отмечены как первые поселенцы лавовых потоков (остров Бали).
Род афананта (Aphananthe) имеет разорванный ареал, охватывающий отдаленные друг от друга районы тропической и теплоумеренной Азии, включая Малезию, районы Соломоновых островов, Восточной Австралии, Мадагаскара и Мексики. Эти огромные дизъюнкции говорят о более широком распространении афананты в прошлом.
В тропиках Южной Азии, Индонезии и Океании (в основном на тропических островах) распространен род жиронъера (Gironniera), представленный огромными вечнозелеными деревьями дождевых тропических лесов (жиронъера каркасолистная — G. ceItidifolia), или небольшими, высотой до 16 м, деревьями более сухих полулистопадных формаций тропиков (жиронъера полуравная — G. snbaequalis и др.).


 Формула и диаграмма цветка
Формула и диаграмма цветка азнообразнейшие биологически активные вещества. Плоды фруктовых и ягодных культур употребляют не только в свежем виде, в огромных масштабах они идут в переработку. Из семян некоторых видов, особенно миндаля, получают пищевые и технические масла, а красивая древесина яблоневых и сливовых идет на различные поделки.
азнообразнейшие биологически активные вещества. Плоды фруктовых и ягодных культур употребляют не только в свежем виде, в огромных масштабах они идут в переработку. Из семян некоторых видов, особенно миндаля, получают пищевые и технические масла, а красивая древесина яблоневых и сливовых идет на различные поделки.





